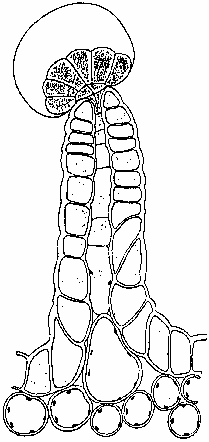
Figure 1. Resin-producing stalked glandular trichome (Briosi and Tognini 1894).
Pate, D.W., 1994. Chemical ecology of Cannabis. Journal of the International Hemp Association 2: 29, 32-37.
The production of cannabinoids and their associated terpenes in Cannabis is subject to environmental influences as well as hereditary determinants. Their biosynthesis occurs in specialized glands populating the surface of all aerial structures of the plant. These compounds apparently serve as defensive agents in a variety of antidessication, antimicrobial, antifeedant and UV-B pigmentation roles. In addition, the more intense ambient UV-B of the tropics, in combination with the UV-B lability of cannabidiol, may have influenced the evolution of an alternative biogenetic route from cannabigerol to tetrahydrocannabinol in some varieties.
Figure 1. Resin-producing stalked glandular trichome (Briosi and Tognini
1894).
Cannabis may have been the first cultivated plant. Records indicate use of this crop for paper, textiles, food and medicine throughout human history (Abel 1980). It is a dioecious annual with rather distinctive palmate leaves, usually composed of an odd number of leaflets. Best growth occurs on recently disturbed sites of high soil nitrogen content, so it is commonly found as a persistent weed at the edge of cultivated fields. Mature height ranges from 1 to 5 meters, according to environmental and hereditary dictates. Typically, the male plant is somewhat taller and more obviously flowered. These flowers have five yellowish tepals, and five anthers that hang pendulously at maturity, dispersing their pollen to the wind. The female plant exhibits a more robust appearance due to its shorter branches and dense growth of leaves and flower-associated bracts. Its double-styled flower possesses only a thin, closely adherent perianth, but is further protected by enclosure in a cuplike bracteole (i.e., perigonal bract), subtended by a usually monophyllous leaflet. A single achene is produced per flower and shed or dispersed as a result of bird predation. The life cycle of the male is completed soon after anthesis, but the female survives until full seed ripeness.
Cannabis seems a virtual factory for the production of secondary metabolic compounds. A variety of alkanes have been identified (Adams, Jr. and Jones 1973, De Zeeuw et al. 1973b, Mobarak et al. 1974a & 1974b), as well as nitrogenous compounds (ElSohly and Turner 1976, Hanus 1975b), flavonoids (Gellert et al. 1974, Paris et al. 1975b, Paris and Paris 1973) and other miscellaneous compounds (Hanus 1976a & 1976b). Terpenes appear in abundance (Hanus 1975a, Hendricks et al. 1975) and contribute to the characteristic odor of the plant (Hood et al. 1973) and some of its crude preparations, such as hashish. The compounds which comprise the active drug ingredients are apparently unique to this genus and are termed cannabinoids. Cannabinoids were originally thought to exist as the phenolic compounds, but later research (Fetterman et al. 1971a, Masoud and Doorenbos 1973, Small and Beckstead 1973, Turner et al. 1973b) has indicated their existence predominantly in the form of carboxylic acids which decarboxylate readily with time (Masoud and Doorenbos 1973, Turner et al. 1973b), upon heating (De Zeeuw et al. 1972a, Kimura and Okamoto 1970) or in alkaline conditions (Grlic and Andrec 1961, Masoud and Doorenboos 1973). There are over 60 of these type compounds present in the plant (Turner et al. 1980).
Much has been published concerning the influence of heredity on cannabinoid production (e.g., Fetterman et al. 1971b, Small and Beckstead 1973), but ecological factors have long been thought to have an important influence by stressing the Cannabis plant (Bouquet 1950). The resultant increased biosynthesis of the cannabinoid and terpene containing resin, in most cases, seems likely of advantage to the organism in adapting it to a variety of survival-threatening situations. This work reviews these biotic and abiotic challenges and speculates on the utility of Cannabis resin to the plant.
The major sites of cannabinoid production appear to be epidermal glands (Fairbairn 1972, Hammond and Mahlberg 1973, Lanyon et al. 1981, Malingre et al. 1975) which exhibit a marked variation in size, shape and population density, depending on the anatomical locale examined. While there are no published reports of glands present on root surfaces, most of the aerial parts possess them, along with non-glandular trichomes (De Pasquale et al. 1974). These epidermal glands seem to fall into two broad categories: stalked and sessile. The stalked gland (Fig. 1, front page) can consist of a single cell or small group of cells arranged in a rosette on a single or multicellular pedestal. Lack of thorough ontogenetic study has led to the speculation that some of this variation may be attributable to observation of various developmental stages (Ledbetter and Krikorian 1975). The sessile gland possesses no stalk and has secretory cells located at or below the epidermal surface (Fairbairn 1972). In either case, the glandular cells are covered with a "sheath" under which the resins are secreted via vesicles (Mahlberg and Kim 1992). This sheath consists of a cuticle that coats a polysaccharide layer (presumed cellulose) originating from the primary cell wall (Hammond and Mahlberg 1978). The resins accumulate until the sheath bulges away from the secretory cells, forming a spheroid structure. The resin is then released by rupture of the membrane or through pores in its surface (De Pasquale 1974). The cannabinoid content of each plant part varies, paralleling observable gland distribution (Fetterman et al. 1971, Honma et al. 1971a & 1971b, Kimura and Okamoto 1970, Ohlsson et al. 1971, Ono et al. 1972), although Turner et al. (1978) have disagreed. Roots contain only trace amounts. Stalks, branches and twigs have greater quantities, although not as much as leaf material. Vegetative leaf contains varying quantities depending on its position on the plant: lower leaves possessing less and upper ones more. Leaf glands are most dense on the abaxial (underside) surface. The greatest amount of cannabinoids is found in the new growth near each apical tip (Kimura and Okamoto 1970, Steinberg et al. 1975), although Ono et al. (1972) seem to differ on this point. This variation in leaf gland placement may be due to either loss of glands as the leaf matures or a greater the endowment of glands on leaves successively produced as the plant matures. Additional study on this point is required.
Once sexual differentiation has occurred, the generation of female reproductive organs and their associated bracts increases total plant cannabinoid content. Bracts subtending the female flowers contain a greater density of glands than the leaves. The small cuplike bracteole (perigonal bract) enclosing the pistil has the highest cannabinoid content of any single plant part (Kimura and Okamoto 1970, Honma et al. 1971a & 1971b). Second only to this is the flower itself (Fetterman et al. 1971b). Since it has no reported epidermal gland structures, the cannabinoids present must be due to either undiscovered production sites or simple adherence of resin from the inner surface of its intimately associated bracteole. This conjecture is supported by the finding that the achenes do not contain substantial amounts of the cannabinoids (Fetterman et al. 1971b, Ono et al. 1972). Reproductive structures of the male plant are also provided with greater concentrations of the cannabinoids (Fetterman et al. 1971b, Ohlsson et al. 1971). Stalked glands have been observed covering the tepal, with massively stalked glands occurring on the stamen filament (Dayanadan and Kaufman 1976). In addition, rows of very large sessile glands are found situated in grooves on the anther itself (Dayanadan and Kaufman 1976, Fairbairn 1972) and apparently provide the pollen with a considerable cannabinoid content (Paris et al. 1975a).
Delta-9-tetrahydrocannabinol (THC) is the cannabinoid responsible for the main psychoactive effects of most Cannabis drug preparations (Mechoulam 1970). In some varieties of Cannabis, additional cannabinoid homologs appear that have the usual pentyl group attached to the aromatic ring, replaced by a propyl (De Zeeuw et al. 1972b & 1973a, Fetterman and Turner 1972, Gill 1971, Gill et al. 1970, Merkus 1971, Vree et al. 1972a, Turner et al. 1973a) or occasionally a methyl group (Vree et al. 1971 & 1972b). Other claims have been made for butyl (Harvey 1976) or heptyl (Isbell 1973) substitutions, but the latter announcement seems particularly tenuous. THC is thought to be produced by the plant (Fig. 2, next page) from cannabidiol (CBD) which, in turn, is derived from cannabigerol (CBG) generated from non-cannabinoid precursors (Hammond and Mahlberg 1994, Shoyama et al. 1984, Turner and Mahlberg 1988). CBG is also the biogenetic precursor of cannabichromene (CBC). Some of the cannabinoids (e.g., cannabielsoin, cannabinol, and cannabicyclol) are probably degradation products of the enzymatically produced cannabinoids (e.g., CBD, THC and CBC, respectively).
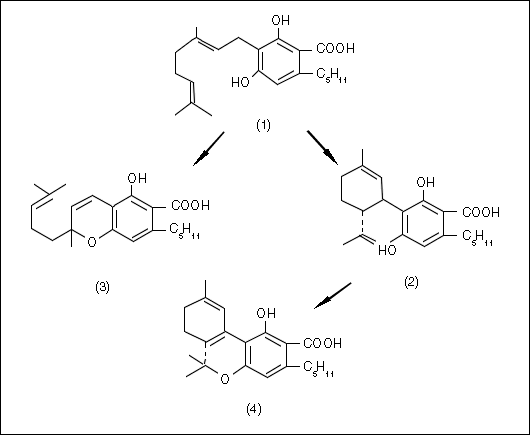
Figure 2. Biosynthesis of cannabinoid acids (redrawn after Shoyama et al. 1975):
1 = cannabigerol (CBG); 2 = cannabidiol (CBD); 3 = cannabichromene (CBC); 4 = delta-9-tetrahydrocannabinol
(THC).
THC is a viscous hydrophobic oil (Garrett and Hunt 1974) that resists crystallization (Gaoni and Mechoulam 1971) and is of low volatility (Adams et al. 1941). Since the sticky resins produced and exuded on the surface of the plant are varying combinations of THC, other cannabinoids and a variety of terpenes, they can be seen as analogous to the waxy coatings of the cacti and other succulents that serve as a barrier to water loss in dry environments.
Bouquet (1950) has mentioned that the western side of Lebanon's mountainous Cannabis growing areas is less favorable for resin production because of humid sea winds. De Faubert Maunder (1976) also observed that the copious separable resin needed for hashish production occurs only "in a belt passing from Morocco eastwards, taking in the Mediterranean area, Arabia, the Indian sub-continent and ending in Indo-China." These are mostly areas notable for their sparse rainfall, low humidity and sunny climate. Is it merely coincidence that resin is produced according to this pattern, as well?
Experimental evidence is accumulating that reinforces these notions. Sharma (1975) reported a greater glandular trichome density on leaves of Cannabis growing in xeric circumstances. Paris et al. (1975a) have demonstrated a marked increase in the cannabinoid content of Cannabis pollen with decreased humidity. Murari et al. (1983) grew a range of Cannabis fiber cultivars in three climatic zones of Italy and found higher THC levels in those plants grown in the drier "continental" (versus "maritime") climate. Hakim et al. (1986) report that CBD-rich English Cannabis devoid of THC produced significant amounts of THC and less CBD, when grown in the Sudan. This trend was accentuated in their next generation of plants.
Haney and Kutscheid (1973) have shown significant correlations of plant cannabinoid content with factors affecting soil moisture availability: content of clay or sand, percent slope of plot, and competition from surrounding vegetation. In some cases, this last factor was noted to have induced a stunted plant with "disproportionally smaller roots", which would tend to increase both the frequency and severity of desiccation stress.
In a study of 10 Kansas locations, Latta and Eaton (1975) found wide differences in plant cannabinoid content, observing that "delta-9-THC ranged from 0.012 to 0.49% and generally increased as locations became less favorable for plant growth, suggesting increased plant stress enhanced delta-9-THC production." Mention was also made of a positive correlation between competing vegetation and THC content. Although the sampling area was not considered very moisture deficient, they speculated that "Greater difference among locations might have been observed under drought conditions."
Temperature may play a role in determining cannabinoid content, but perhaps only through its association with moisture availability. Boucher et al. (1974) reported an increase in cannabinoid content with temperature (32o C. vs. 22o C.), however, some variables such as increased water loss due to accelerated evaporation and plant transpiration at high temperatures were left unaccounted. In contrast, Bazzaz et al. (1975), using 4 Cannabis ecotypes of both tropical and temperate character, demonstrated a definite decrease in cannabinoid production with increased temperature (32o C. vs. 23o C.). Later studies by Braut-Boucher (1980) on clones of 2 strains from South Africa revealed a more complex pattern of biosynthesis according to strain, gender and chemical homologue produced. Clearly, further study of this parameter is needed.
Mineral balance seems to influence cannabinoid production. Krejci (1970) found increases related to unspecified "poor soil conditions". Haney and Kutcheid (1973) have shown the influence of soil K, P, Ca and N concentrations on Illinois Cannabis. They report a distinctly negative correlation between soil K and plant delta-9-THC content, although K-P interaction, N and Ca were positively correlated with it. These minerals were also shown to affect the production of CBD, delta-8-THC and cannabinol (CBN), although the latter two compounds are now thought to be spontaneous degradation products of delta-9-THC. Kaneshima et al. (1973) have demonstrated the importance of optimal Fe levels for plant synthesis of THC. Latta and Eaton (1975) reported Mg and Fe to be important for THC production, suggesting that these minerals may serve as enzyme co-factors. Coffman and Gentner (1975) also corroborated the importance of soil type and mineral content, and observed a significant negative correlation between plant height at harvest and THC levels. Interestingly, Marshman et al. (1976) report greater amounts of THC in Jamaican plants growing in "organically" enriched (vs. artificially fertilized) soils.
Wounding of the plant has been employed as a method to increase resin production (Emboden 1972). This increase may be a response to desiccation above the point of vascular disruption. Under natural circumstances, wounding most often occurs as a result of insect attack. This is a source of environmental stress which the production of terpenes and cannabinoids may be able to minimize. Cannabis is subject to few predators (Smith and Haney 1973, Stannard et al. 1970) and has even been utilized in powdered or extract form as an insecticide (Bouquet 1950) or repellent (Khare et al. 1974). Its apparent defensive mechanisms include a generous covering of non-glandular trichomes, emission of volatile terpenoid substances, and exudation of the sticky cannabinoids. Cannabis is often noted for its aromatic quality and many of the terpenes produced are known to possess insect-repellent properties. Among these are alpha and beta pinene, limonene, terpineol and borneol. Pinenes and limonene comprise over 75% of the volatiles detected in the surrounding atmosphere, but account for only 7% of the essential oil (Hood et al. 1973). Consistent with glandular trichome density and cannabinoid content, more of these terpenes are produced by the inflorescences than the leaves, and their occurrence is also greater in the female plant (Martin et al. 1961).
No insect toxicity studies using isolated cannabinoids have been published to date. Rothschild et al. (1977) found THC-rich Mexican (vs. CBD-rich Turkish) Cannabis fatal to tiger moth (Arctia caja) larvae, but not Nigerian grasshopper (Zonocerus elegans) nymphs. Rothschild and Fairbairn (1980) later found that pure THC (vs. CBD) sprayed on cabbage leaves, does repel the large white cabbage butterfly (Pieris brassicae).
The cannabinoids may also serve as a purely mechanical defense. A tiny creature crossing the leaf surface could rupture the tenuously attached globular resin reservoirs of the glandular trichomes (Ledbetter and Krikorian 1975) and become ensnared in resin. A sizable chewing insect, if able to overcome these defenses, would still have difficulty chewing the gummy resin, along with the cystolithic trichomes and silicified covering trichomes also present on the leaf. The utility of these epidermal features as insect antifeedants is also inferable from their predominant occurrence on the insect-favored abaxial leaf surface. Although the above strategies represent a seemingly sophisticated system, many other plants (Levin 1973) and even arthropods (Eisner 1970) utilize similar defense mechanisms, often employing identical terpenes!
Terpenes may also help to suppress the growth of surrounding vegetation (Muller and Hauge 1967, Muller et al. 1964). Haney and Bazzaz (1970) speculated that such a mechanism may be operative in Cannabis. They further ventured that since the production of terpenes is not fully developed in very young plants, this may explain their inability to compete successfully with other vegetation until more mature. The observation (Latta and Eaton 1975) of increased THC production by plants in competition with surrounding vegetation "at a time in the growing season when moisture was not limiting", may indicate a stimulus for cannabinoid production beyond that of simple water stress.
The cannabinoids may serve as a protectant against microorganisms. Cannabis preparations have long served as medicines (apart from their psychoactive properties) and are effective against a wide variety of infectious diseases (Kabelic et al. 1960, Mikuriya 1969). These antibiotic properties have been demonstrated with both Cannabis extracts (Ferenczy et al. 1958, Kabelic et al. 1960, Radosevic et al. 1962) and a variety of isolated cannabinoids (ElSohly et al. 1982, Farkas and Andrassy 1976, Gal and Vajda 1970, Van Klingeren and Ten Ham 1976). CBG has been compared (Mechoulam and Gaoni 1965) in both "structure and antibacterial properties to grifolin, an antibiotic from the basidiomycete Grifolia conflens." Ferency (1956) has demonstrated the antibiotic properties of Cannabis seed, a factor that may aid its survival when overwintering. Adherent resin on the seed surface, as well as a surrounding mulch of spent Cannabis leaves, may serve in this regard.
Some of the many fungal pathogens that affect Cannabis include Alternaria alterata (Haney and Kutsheid 1975), Ascochyta prasadii (Shukla and Pathak 1967), Botryosphaeria marconii (Charles and Jenkins 1914), Cercospora cannabina and C. cannabis (Lentz et al. 1974), Fusarium oxysporum (McCain and Noviello 1985), Phoma sp. (Srivastava and Naithani 1979) and Phomopsis ganjae (McPartland 1984).
While A. alterata attacks Illinois Cannabis and destroys 2.8-45.5% of the seed (Haney and Kutsheid 1975), the balance of these species are leaf spot diseases. McPartland (1984) has demonstrated the inhibitory effects of THC and CBD on Phomopsis ganjae. However, De Meijer et al. (1992), in evaluating a large collection of Cannabis genotypes, did not find a correlation between cannabinoid content and the occurence of Botrytis. Fungal evolution of a mechanism for overcoming the plant's cannabinoid defenses may be responsible for their success as pathogens. Indeed, some have been demonstrated to metabolize THC and other cannabinoids (Binder 1976, Binder and Popp 1980, Robertson et al. 1975).
Another stress to which plants are subject results from their daily exposure to sunlight. While necessary to sustain photosynthesis, natural light contains biologically destructive ultraviolet radiation. This selective pressure has apparently affected the evolution of certain defenses, among them, a chemical screening functionally analogous to the pigmentation of human skin. A preliminary investigation (Pate 1983) indicated that, in areas of high ultraviolet radiation exposure, the UV-B (280-315 nm) absorption properties of THC may have conferred an evolutionary advantage to Cannabis capable of greater production of this compound from biogenetic precursor CBD. The extent to which this production is also influenced by environmental UV-B induced stress has been experimentally determined by Lydon et al. (1987). Their experiments demonstrate that under conditions of high UV-B exposure, drug-type Cannabis produces significantly greater quantities of THC. They have also demonstrated the chemical lability of CBD upon exposure to UV-B (Lydon and Teramura 1987), in contrast to the stability of THC and CBC. However, studies by Brenneisen (1984) have shown only a minor difference in UV-B absorption between THC and CBD, and the absorptive properties of CBC proved considerably greater than either. Perhaps the relationship between the cannabinoids and UV-B is not so direct as first supposed. Two other explanations must now be considered. Even if CBD absorbs on par with THC, in areas of high ambient UV-B, the former compound may be more rapidly degraded. This could lower the availability of CBD present or render it the less energetically efficient compound to produce by the plant. Alternatively, the greater UV-B absorbency of CBC compared to THC and the relative stability of CBC compared to CBD might nominate this compound as the protective screening substance. The presence of large amounts of THC would then have to be explained as merely an accumulated storage compound at the end of the enzyme-mediated cannabinoid pathway. However, further work is required to resolve the fact that Lydon's (1985) experiments did not show a commensurate increase in CBC production with increased UV-B exposure.
This CBC pigmentation hypothesis would imply the development of an alternative to the accepted biochemical pathway from CBG to THC via CBD. Until 1973 (Turner and Hadley 1973), separation of CBD and CBC by gas chromatography was difficult to accomplish, so that many peaks identified as CBD in the preceding literature may in fact have been CBC. Indeed, it has been noted (De Faubert Maunder 1970) and corroborated by GC/MS (Turner and Hadley 1973) that some tropical drug strains of Cannabis do not contain any CBD at all, yet have an abundance of THC. This phenomenon has not been observed for northern temperate varieties of Cannabis. Absence of CBD has led some authors (De Faubert Maunder 1970, Turner and Hadley 1973) to speculate that another biogenetic route to THC is involved. Facts scattered through the literature do indeed indicate a possible alternative. Holley et al. (1975) have shown that Mississippi-grown plants contain a considerable content of CBC, often in excess of the CBD present. In some examples, either CBD or CBC was absent, but in no case were plants devoid of both. Their analysis of material grown in Mexico and Costa Rica served to accentuate this trend. Only one example actually grown in their respective countries revealed the presence of any CBD, although appreciable quantities of CBC were found. The reverse seemed true as well. Seed from Mexican material devoid of CBD was planted in Mississippi and produced plants containing CBD.
Could CBC be involved in an alternate biogenetic route to THC? Yagen and Mechoulam (1969) have synthesized THC (albeit in low yield) directly from CBC. The method used was similar to the acid catalyzed cyclization of CBD to THC (Gaoni and Mechoulam 1966). Reaction by-products included cannabicyclol, delta-8-THC and delta-4,8-iso-THC, all products which have been found in analyses of Cannabis (e.g., Novotny et al. 1976). Finally, radioisotope tracer studies (Shoyama et al. 1975) have uncovered the intriguing fact that radiolabeled CBG fed to a very low THC-producing strain of Cannabis is found as CBD, but when fed to high THC-producing plants, appeared only as CBC and THC. Labeled CBD fed to a Mexican example of these latter plants likewise appeared as THC. Unfortunately, radiolabeled CBC was not fed to their plants, apparently in the belief that CBC branched off the biogenetic pathway at CBD and dead ended. Their research indicated that incorporation of labeled CBG into CBD or CBC was age dependent. Vogelman et al. (1988) likewise report that the developmental stage of seedlings, as well as their exposure to light, affects the occurrence of CBG, CBC or THC in Mexican Cannabis. No CBD was reported.
Although the chemistry of Cannabis has come under extensive investigation, more work is needed to probe the relationship of its resin to biotic and abiotic factors in the environment. Glandular trichomes are production sites for the bulk of secondary compounds present. It is probable that the cannabinoids and associated terpenes serve as defensive agents in a variety of antidessication, antimicrobial, antifeedant and UV-B pigmentation roles. UV-B selection pressures seem responsible for the distribution of THC-rich Cannabis varieties in areas of high ambient radiation, and may have influenced the evolution of an alternate biogenetic pathway from CBG to THC in some of these strains. Though environmental stresses appear to be a direct stimulus for enhanced chemical production by individual plants, it must be cautioned that such stresses may also skew data by hastening development of the highly glandular flowering structures. Future studies will require careful and representative sampling to assure meaningful results.