80
Physical evidence for the antiquity of Cannabis sativa L.
Michael P. Fleming1 and Robert C. Clarke2
1 Brooklyn
Botanic Garden, 100 Washington Ave., Brooklyn, NY, USA 11225-1099
Tel: +1 (718) 941-4044 ext. 219, Fax: +1 (718) 941-4774; e-mail: michaelfleming@mail.bbg.org
2 International
Hemp Association, Postbus 75007, 1070 AA Amsterdam, The Netherlands
Tel/Fax: +31 (0)20 618-8758; e-mail: iha@euronet.nl
Fleming, M. P. and R. C. Clarke 1998. Physical evidence for the antiquity of Cannabis sativa L. (Cannabaceae). Journal of the International Hemp Association 5(2): 80-92. Cannabis has been an important economic crop plant for six millennia. Its uses for fiber, food, oil, medicine, and as a recreational/religious drug have been prevalent throughout this period. Recent palynological research into the agricultural and environmental history of Cannabis has produced curves for Cannabaceae pollen at a number of sites in Europe and America. Additional archaeological remains and written records provide evidence for both Old and New World occurrences. This paper discusses the origin, domestication and migration of hemp as a crop plant as documented by palynological and archaeological evidence. In addition, the comparative morphology of Cannabis and Humulus pollen grains is described, and the problems of interpreting Cannabaceae pollen in the stratigraphic record are discussed.
Introduction
Since prehistoric times, Cannabis
has been with us in one form or another. As an adaptable weedy annual, it has,
under human tutelage, successfully extended its range, both latitudinally and
longitudinally (Li 1974). Paleobotany, and in particular palynology, offer the
most accurate evidence of the growth of a plant in a particular region, even
though plant remains can be carried by water and wind over great distances
before they are deposited. Several types of paleobotanical physical evidence of Cannabis,
such as pollen grains, fibers, seeds, fiber and seed impressions, trichomes,
carbonized remains and cannabinoid compounds, have been recovered from dated
archeological contexts. Pollen evidence has proven very useful in establishing
the early range of Cannabis in Europe. Cord and textile scraps as well as
fiber impressions in pottery and bronze figures have been cited as early
evidence of hemp in Asia. Seed remains have been recovered from both Europe and
Asia. Cannabis trichomes (plant hairs) are resistant to decay and burning
and are also occasionally recovered. Carbonized remains are relatively rare and
are confined to eastern Africa and southern Europe. Cannabinoid compounds unique
to the genus Cannabis are also occasionally identified. These various
types of physical evidence allow us to substantiate or refute historical records
concerning the origin and early diffusion of Cannabis. The various
classes of physical evidence are discussed below in more detail.
Pollen
Hemp (Cannabis sativa
L.) and hop (Humulus spp.) had previously been classified into the
Moraceae and the Urticaceae, but are now widely accepted as belonging to their
own family, the Cannabaceae (sometimes incorrectly called the Cannabinacaeae).
The taxonomy and classification of the Cannabaceae have always been disputed (Schultes
1970), but current classification schemes recognize only one species Cannabis
sativa L. with various subspecies and varieties (Chrtek 1981, Small and
Cronquist 1976), three species of Humulus (H. lupulus, H. japonicus
and H. yunnanensis) (Small 1978), and a recently described new genus and
species, Humulopsis scandens (Lour.) I. A. Grudzinskaya (Grudzinskaya
1988). Cannabaceae pollen grains are most often recovered in sediments from the
bottom of lakes, ponds and wells.
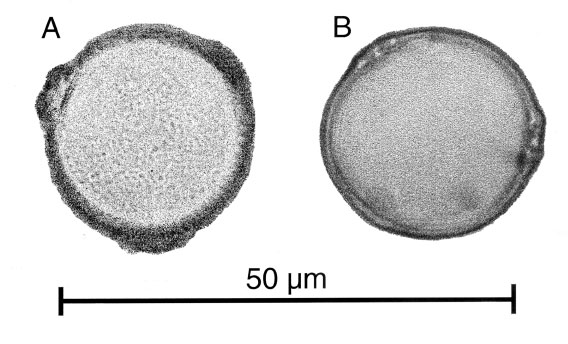
The pollen grains of the family have
been sketched and described by several authors (Walker 1955, Godwin 1967a,
French and Moore 1986, Whittington and Gordon 1987, Whittington and Edwards
1989). Cannabaceae pollen consists of trizonoporate grains typically less than
50 microns in diameter. The pore complex is generally not greater than 3-4
microns in diameter.
French and Moore (1986) described
Cannabaceae pollen in detail, noting that,
"...at the break of the slope the sexine and nexine separate as if to form a vestibulum. The nexine continues to follow the circumference of the grain for approximately 0.5 microns. In some grains it is seen to taper out, whilst in many grains it is not discernible. The tectum rises away from the nexine to form the pore annulus before steeply penetrating below the level of the endexine. The diameter of the pore cavity is 1.0 to 1.5 microns and is often slightly larger at the base. Underneath the annulus is a void, sometimes referred to as the hollow internal annulus. Away from the pore the exine is thin."
Due to the close
resemblance between Cannabis and Humulus pollen grains, the
similar flowering times for both genera, and the fact that both genera shed
buoyant pollen in vast quantities (Lewis et al. 1983) there have been
difficulties in accurately interpreting the Cannabaceae pollen records from
past site analyses. The desirability of separating these two species has
encouraged researchers to designate several characteristics which can be used to
determine whether a Cannabaceae pollen grain came from a hemp or hop plant
(Whittington and Edwards 1989). Godwin (1967a) examined the pore complexes of Cannabis
and Humulus pollen and noted differentiating factors between the two,
based on several minute details in these complexes. Described later in
independent studies by French and Moore (1986) and Whittington and Gordon
(1987), these factors include the grouping of the scabrae as revealed by
scanning electron microscope (SEM), the virtual absence of the hollow internal
annulus in hop grains, the rise and arch of the tectum over the rim of the
pores, and the steep slope of the annulus of Cannabis versus the low
slope of Humulus pore complexes.
Godwins criterion of whether or
not the tectum penetrates beneath the endexine was judged too cumber-some.
French and Moore (1986), Whittington and Edwards (1989), Whittington and Gordon
(1987) and Paahlson (1981) have all concluded that statistically significant
values for separation can be achieved by noting pollen grain diameter and the
degree of pore uplift. Of course, these methods may be supplemented by observing
the aforementioned details of the pore complexes. French and Moore (1986) noted
that greater than 60% of Cannabaceae pollen can be differentiated by the degree
of pore protrusion alone, noting that the pore protrudes proportionately much
more on Cannabis grains than on those of hop. However, their calculations
were based on a small sample size, and some of their samples were distorted by
size increases in the grains due to prolonged acetolysis and storage in glycerol
jelly. Whittington and Gordon (1987) reduced the time of acetolysis and
substituted silicone oil for glycerol. Their data showed that by noting pore
protrusion and pollen grain diameter, one can estimate the proportion of Humulus
and Cannabis grains in a mixed sample. Their study was based on the
assumption that "the proportion of pollen grains falling into each category
of the pure populations are assumed to be relevant figures for the analysis of
mixtures of pollen grains." Whittington and Edwards (1989) also addressed
the problem of hemp and hop pollen differentiation based on pollen diameter and
pore protrusion. They showed that while the pollen of Cannabis is
generally larger than that of Humulus, this in itself was an unreliable
criterion for separation and must be supplemented with data on pore complex
protrusion.
These conclusions do not necessarily
indicate that all previous attempts at interpreting Cannabaceae pollen curves
and separating the two genera are incorrect, but it does mean that less
confidence can be attached to earlier studies. It should also be remembered that
pore protrusion and pollen grain size methods of separation are
based on statistical methods and are dependent on a sufficiently large
population of measurable grains. The tendency for palynologists to ascribe a
marked expansion of the Cannabaceae curve to Cannabis sativa may
be reasonable and correct, but local occurrences of Humulus lupulus
may mean that the presence or importance of hop is being underestimated. Also,
it is important to consider that Cannabis produces copious amounts of
pollen, where Humulus tends to shed relatively much less pollen (Lewis et
al. 1983). According to Whittington and Edwards (1989):
"Given the notoriety which surrounds the present day use of Cannabis crops in drug production, it seems unlikely that pollen trapping experiments from Europe and North America can be carried out on the scale necessary to provide convincing analogues for past patterns of production".
All the ambiguity
interpreting the Cannabis pollen record also means that it is not yet
possible to suggest that a certain proportion of Cannabis pollen would
indicate on-site retting (i.e., the process in which stems are soaked in
water for prolonged periods to free their bast fibers) rather than, or along
with, local cultivation. Cannabis in the fossil pollen assemblages
recovered from lake sediments may come from locally growing hemp whose pollen
has been aerially transported to the lake or from hemp of local or foreign
origin transported to the lake for retting. The existence of known retting
sites, however, are certainly worthy of more palynological investigation. The
pollen method could also be used to verify the use of such sites for hemp
retting purposes. Finally, awareness on the part of analysts that hop or flax (Linum),
another major crop plant utilized for its fibers, that may have been locally
cultivated or retted prior to, during, or after Cannabis, may
produce more comprehensive data and would be of both palynological and
historical interest (Whittington and Edwards 1989).
Flax may be underrepresented in the
pollen record because it is insect pollinated rather than wind pollinated and
therefore it does not produce as much free pollen as either hop or hemp.
However, flax pollen does show up in spikes concurrent with other signs of human
settlement (such as charcoal layers signifying clearing of the land) and the
cultivation of other indicator species such as cereals.
Figure 1. Pollen grains of (A): Cannabis sativa L and (B): Humulus lupulus L. Modified from photos taken by Keith Bennett. Used with permission.
Fibers
Schultes and Hofmann (1992)
provided an overview of macrofossil evidence of the migration of Cannabis
through the Old World. Fiber remains of Cannabis are usually found as
scraps of cordage, textile fragments or pieces of paper. Mats of loose fibers
have also been recovered from lake sediments in Europe. The vast majority of Cannabis
fiber remains have been recovered from sites in China. The identity of fibers as
Cannabis is often based on the context in which the fiber remains were
found. Although it is possible to differentiate commercially available hemp
fiber from other bast fibers by microscope techniques and chemical analysis,
there is still much confusion surrounding the positive identification of
degraded fibers recovered from archeological sites. In almost all cases, no
actual laboratory identification of the plant fibers is provided and they are
called "hemp" largely because of their context in early Chinese
remains. If these same fiber samples had been recovered in western Europe rather
than China, they might very well have been assumed to be flax rather than hemp,
as flax also had a long history in ancient Europe.
Well preserved cord and fabric
samples recovered from several important excavations may be made of hemp, but no
definitive fiber identity has been reported in the literature. What fiber was
used to twist the cord skirt of the Danish Bronze Age (ca. 3250 BP)
Egtved girl or the rope noose around the neck of the well preserved Tollund Mans
body recovered from a Danish bog? What fiber was used to weave the burial
textiles of the Celtic chief buried at Hochdorf (ca. 2550-2500 BP), the
tunics of the Iron Age (ca. 2400 BP) Pazyryk nobles and Ukok Princess (Bahn
1996) or the hunting nets of early (ca. 26,980 to 24,870 BP) Gravettians
(Pringle 1977). It would be interesting to examine these archeological remains
with more modern analysis techniques in order to accurately determine fiber
identity.

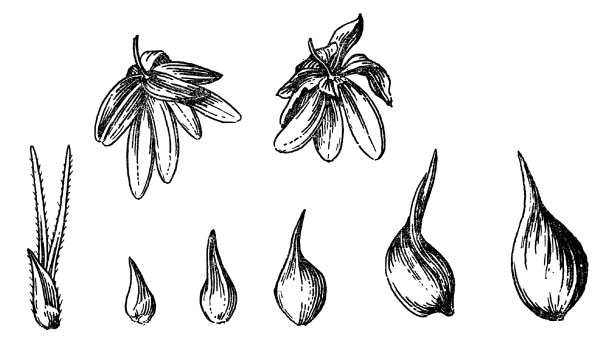
Figure 2. Modern 'Novosadska' hemp, Pazyryk Iron Age and British medieval Cannabis seeds.
Seeds
Cannabis achene fruits,
commonly known as seeds, have been found at many sites. Cannabis seeds
are much easier to positively identify in archeological contexts than pollen
grains or fibers. The characteristic shape of Cannabis seeds is not
easily confused with other genera. Even when partial seeds are recovered, they
often are half seeds and can still be identified (Figure 2). Dörfler (1990)
published a review article of Cannabis seed and pollen remains from
continental Europe, where the majority of Cannabis seed remains have been
recovered. Cannabis seed remains are often considered indicative of hemp
retting, especially in the presence of Cannabaceae pollen. Gaillard (pers. comm.
1997) interpreted high amounts of Cannabis pollen along with only a few
seeds in retting ponds as evidence that predominantly male (pollen) plants were
used for fiber production rather than female (seed) plants.
Fiber and seed impressions
Impressions of plant parts, such as
fibers and seeds, are often recovered from archeological sites. Fiber
impressions attributed to Cannabis have been found in earthen floors and
clay pottery and as well as in the corroded surface of metal objects. Plant
fiber impressions most often appear as cord marks either intentionally pressed
into the exposed surface of pottery as a decoration, or as cord or textile
patterns in the bottom of pottery, resulting from drying it on a woven cloth or
cordage mat. Another type of textile impression is found in the corroded surface
of bronze artifacts that were wrapped in cloth.
Fiber impressions in clay pottery,
soil and metal objects present only circumstantial evidence for the antiquity of
Cannabis (or other plant genera), because the fibers themselves are
subject to decay. The context in which the fiber impressions are recovered is
very important in deciding the identity of the plant fiber represented by the
impressions. Even when cordage and textile imprints are found along with Cannabis
seed or pollen remains, indicating that Cannabis grew nearby and was used
for food, the imprints do not necessarily mean that they were made by hemp
fibers.
Barber (1991) points out that,
"Archeologically, the earliest known candidates for hempen cloth occur in the form of impressions on East Asian Neolithic pots . . .. Although it is not possible to prove directly that the fiber that made these impressions was hemp, the circumstantial evidence is strong for northern China. The fibers are too coarse to have been silk; no other fiber-producing source was known to have existed in northern China until the much later introduction of ramie and cotton from the south; and fair quantities of what is probably hemp pollen have been found at Pan-po; one of the sites with these early textile impressions (Li 1974b)."
Pringle (1997), reporting on Gravettian sites in the Czech Republic dated at ca. 26,980 to 24,870 BP, refers to impressions of plant fiber cordage in clay fragments from the floors of dwellings,
"Adovasio, one of the worlds experts on prehistoric fiber technology, quickly recognized the imprints of basketry or textiles on four fragments . . .. Almost certainly, says Adovasio, the impressions were created from fabrics woven of fibers from wild plants, such as nettle or wild hemp, that were preserved by accident."
"Hyland [another researcher investigating the remains] also discovered impressions of cordage ranging in diameter from 0.31 to 1.15 millimeters and bearing weavers knots, a technique for joining two lengths of cords that is commonly used to make nets of secure mesh."
The estimated mean mesh diameter was 4 mm. and the nets were
probably used for trapping birds and small game. If these impressions could be
positively identified as those of Cannabis they would be by far the
oldest archeological evidence for Cannabis antiquity. However, without
further substantiation, the researchers are only speculating.
We also consider impressions of seeds
to be circumstantial evidence, if the actual seed no longer remains. However,
the shape and size of the Cannabis achene is more distinctive than the
impression left by a bundle of plant fibers, and is easier to identify.
Carbonized and chemical remains
The distinctive shape of Cannabis
trichomes (hairs) allows their identification even in semi-combusted material (Nordal
1970) In addition, carbonized remains can be analyzed for the presence of the
stable cannabinoid Δ6-tetrahydrocannabinol
(Δ6-THC = Δ8-THC)
a compound unique to Cannabis. The presence of Δ6-THC
results from the acid catalysis of Δ1-THC
(Δ1-THC = Δ9-THC)
and cannabidiol (CBD) to Δ6-THC during the
burning process (Zias et al. 1993). In certain instances, cannabinoid
com-pounds can also be isolated from non-carbonized remains.
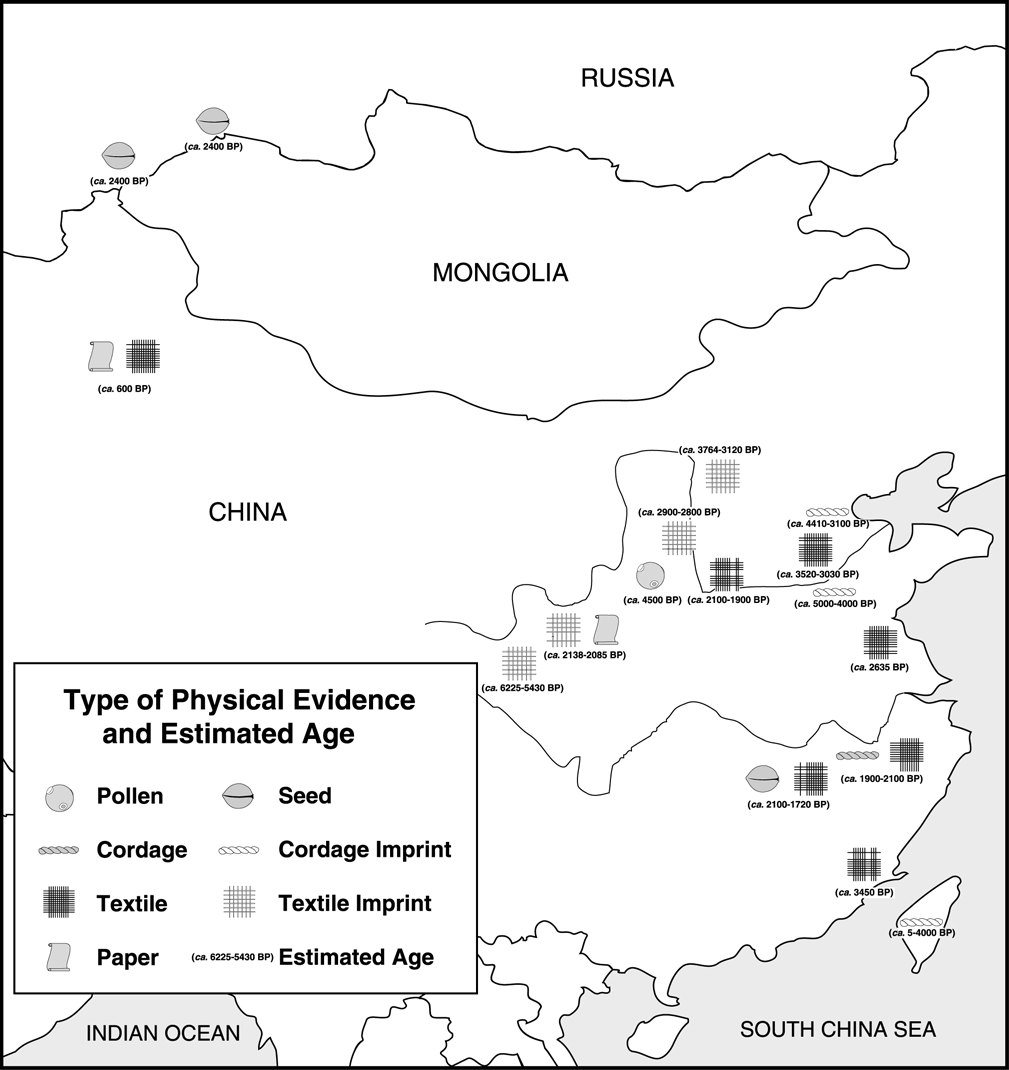
Figure 3. Asian Cannabis remains and approximate ages.
Physical evidence for the origin and diffusion of Cannabis in Asia
"Many cultivated plants are so changed from their ancestral types that it is not possible to unravel their evolutionary history. Such is not the case, however, with Cannabis. Yet, despite its long history as a major crop plant, Cannabis is still characterized more by what is not known about its biology than what is known (Schultes and Hofmann 1992)."
This situation, however, is changing. Work
by Vavilov (1987) has shed light on how Cannabis may have been originally
domesticated. It is well known that Cannabis requires soil with a high
nutrient content, either artificially fertilized or naturally occurring. By
working with wild hemp growing in Mongolia, Vavilov imitated the process of
selection and domestication of hemp as it may well have occurred 6500 years ago.
Vavilov postulated four stages in the
domestication of Cannabis: (1) existence of plants entirely in their wild
state, (2) initial colonization of the wild plant on nutrient-rich dump heaps,
(3) utilization of the weed by local inhabitants, and (4) intentional
cultivation (Vavilov 1992). Unlike oats and rye, which required an intentional
effort to locate and utilize, hemp was very likely only circumstantially
domesticated. Domestication probably occurred independently in several centers
in northeast Asia around six millennia ago (Vavilov 1987, Schultes 1970, Li 1973
and 1974).
Due to biological data in English
translation being recently released from China and India (Harlan 1986), details
of the centers and exact dates of Cannabis domestication vary. We were
able to locate little palynological data for the Asian continent (Figure 3).
Only Chou (1963) reveals a pollen curve for what he interpreted as Humulus
at Pan-po, China dated to ca. 4500 BP. Recently, Garg (1996) has found
Cannabis pollen in the pollen loads of bees in present-day northeastern
India. As a result, we rely largely on archaeological evidence. It is widely
accepted that the Chinese were the first to domesticate this native Asian plant.
There is strong archaeological evidence of its widespread use as an economic
crop by ca. 6500 BP. Existing records place its major center of
domestication in present-day northern China where there is a continuous record
of its use from the Neolithic to the present (Li 1973). While its use as a food
source probably resulted in its initial domestication, Cannabis fiber was
discovered not long thereafter.
In ancient China, hemp cords were
used to form the core of clay temple statues. Remains of Song Dynasty (ca.
1040 to 720 BP) statuary made with hemp cordage have been excavated near Chin-Cheng
in Shanxi Province (Kao 1978). Hemp textile, rope and thread remains were also
reported from a Liang-Chu culture site in Zhejiang Province (Cheng 1966). Hemp
textile remains have been discovered at a Shan Kingdom (ca. 3520 to 3030 BP)
site in Anyang in Henan Province (Chang 1963). Zhou Dynasty (ca. 3200 to
2219 BP) cemeteries in Anyang, and also Changsha in Hunan Province, yielded
thousands of funerary objects, and the inventories listed included hemp textiles
(Cheng 1963). A Zhou Dynasty tomb in Shanxi Province contained hemp cloth of a
light weave, indicating that hemp weaving had reached a fairly high standard (Li
1974). Excavations of Han Dynasty (ca. 2100 to 1900 BP) tombs in Gansu
Province produced complete specimens of hemp cloth used to cover corpses. The
coverings were wrapped around silk dresses and were tied with hemp ropes. Hemp
fibers were also used for reinforcing the plaster covering of the brick walls of
the crypt (Kansu Museum 1972). Laquerware winged cups constructed over hemp
cloth cores were recovered from a Han Dynasty (ca. 2100 to 1900 BP)
wooden pit burial near Nanchang city, Jiangxi Province (Kuo 1978). A boat coffin
recovered from the Wuyi mountains in Fujian Province and carved from a single
tree trunk contained the body of an old man wrapped in cloth funeral shroud said
to be made of jute, hemp, silk and cotton. The coffin was dated at ca.
3600 to 3300 BP (Li 1984). Fragments of both silk and hemp textiles dated to ca.
2655 to 2615 BP were found in a tomb excavated in Anhui Province adhering to the
outer surface of bronze sculptures indicating that the bronzes had been wrapped
in cloth (Yin 1978). Many other textile samples identified as "hemp"
have been recovered from additional sites across China, but these are undated
and have been omitted from this survey.
China is the original home of paper
making, and paper scraps containing hemp fibers have been discovered at Chinese
archeological sites. Li (1974) and Temple (1988) both report on the 1957
discovery of a scrap of paper in a tomb near Xian in Shaanxi Province dated
at ca. 2138 to 2085 BP. This is considered to be the oldest surviving
piece of paper ever recovered from a dated site. Temple (1988) describes the
piece of paper and explains its fabrication,
"It is about 10 cm square and can be dated precisely between the years 140 and 87 BC [ca. 2138 to 2085 BP]. This paper and similar bits of paper surviving from the next century are thick, coarse and uneven in their texture. They are all made of pounded and disintegrated hemp fibers. From the drying marks on them, it is evident that they were dried primitively on mats woven as pieces of fabric [also likely hemp], not on what we know as paper molds. In these early days, the water just drained slowly through the underlying mat of fabric, leaving the paper layer on top. This was then peeled off and dried thoroughly. But so thick and coarse was the result, that it could not have been very satisfactory for writing."
Li (1974) describes a much later find of hemp paper along with other hempen artifacts at Turfan in Xinjiang Province in western China,
"In one grave was found a rare fragmentary script of the Lun Yü (Analects of Confucius) written in 716 AD [ca. 1282 BP] on white hemp paper. Also found were paper shoes made of pasted layers of white hemp paper sewn together with white hemp threads. In the same grave was a complete cloth sheet of hemp fabric.
"In another grave dated 721 AD [ca. 1277 BP], there was hemp cloth as well as hemp shoes. The latter were of two kinds, one woven of hemp fibers and the other sewn in a fine yellow cloth."
China has produced fewer seed remains than fiber remains and fiber impressions. Li (1974b) describes the tomb of a woman from Han Dynasty Tomb No. 2 (ca. 2100 to 1900 BP) at Changsha, Hunan Province,
"Besides fruits such as pears, peaches and jujubes, there were grains such as rice, wheat, millet, hemp seed, and mustard seed. Hemp seed was clearly used in early Han times as a common grain along with the other cereals."
The most well publicized, as well as the most controversial, Cannabis seed remains (Figure 2) were recovered from the frozen tomb of Iron Age (ca. 2430 BP) nobles of eastern Siberia discovered by Sergei Rudenko in 1929 (Rudenko 1970). We have included Rudenkos account of the occurrence of Cannabis seeds in the tomb, even though much of it is highly conjectural.
"Thus in barrow 2, two smoking sets were found: vessels containing stones that had been in the fire and hemp seeds; above them were shelters supported on six rods, in one case covered with a leather hanging and in the other case probably with a felt hanging, large pieces of which were found in the southwest corner of the tomb. Finally, there was a [leather] flask containing hemp seeds [Figure 2] fixed to one of the legs of a hexapod stand. Consequently we have the full set of articles for carrying out the purification ritual, about which Herodotus wrote in such detail in his description of the Black Sea Scyths. There had been sets for smoking hemp in all the Pazyryk barrows; the sticks for the stand survived in each barrow although the censers and cloth covers had all been stolen except in barrow 2. Hemp smoking was practiced evidently not only for purification, but in ordinary life by both men and women.
"In each vessel besides the stones, as already mentioned above, there was a small quantity of seeds of hemp (Cannabis sativa L. of the variety C. ruderalis Janisch.). Burning hot stones had been placed in the censer and part of the hemp seeds had been charred. Furthermore the handle of the cauldron censer had been bound round with birch bark, evidently because the heat of the stones was such that its handle had become too hot to hold in the bare hands."
Rudenkos
speculations on the smoking of Cannabis by these Iron Age nobles, based
largely on Herodotus accounts of the Scythians of the Black Sea region, have
led to many claims that the Scythians smoked Cannabis. Clarke (1998)
provides a more complete rebuttal of these claims. All that we know with
certainty is that Cannabis seeds were found in the Pazyryk tombs. Woven
textiles were also recovered, but the identity of the fibers has yet to be
determined (Clarke 1995).
Cannabis remains may also
have been recovered from more recently excavated Iron Age tombs similar to the
Pazyryk tombs discovered by Rudenko. In 1993, a tomb containing the preserved
body of a young woman nicknamed "The Lady" or the "Frozen
Princess" was discovered by a Russian team headed by Natalia Polosmak. The
tomb dated at ca. 2400 BP reportedly contained many grave goods,
including either Cannabis remains in a small pot (Anon. 1994) or
coriander seeds in a stone dish (Polosmak 1996). It is not clear if the seed
remains were of Cannabis or coriander or both. Polosmak (1996) also
echoes Rudenkos (1970) theory that the Pazyryk people may have breathed the
fumes of burning Cannabis.
Nearly all of the remaining early
fiber impressions attributed to Cannabis hemp originate from China. Li
(1974b) mentions several sites in China where cordage impressions deduced as
being Cannabis were recovered from various sites of the Yang-Shao culture
in northern China dated ca. 4410 to 3100 BP and from a late Neolithic age
(ca. 5000 to 4000 BP) site in Henan Province in eastern China.
Additional evidence of cord
impressions have been recovered from early Neolithic (ca. 6000 to 5000
BP) sites on Taiwan island. According to Cheng (1959), referring to the lower
stratum of Yuan Shan, the richest and most important site on the island,
"The characteristic remains here as elsewhere on the island, are the cord-marked pottery, a coarse sandy ware, handmade, thick-walled, with a plain surface covered with cord marks and, occasionally, with lineal impressions. . . . There is also a stone beater, a rod-shaped implement with incisions, which might have been used for the lineal impressions in pottery decoration, or for pounding hemp fiber, a common raw material for rope and textile."
Textile imprints
have been found in pottery shards, clay layers and bronze objects. Anderson
(1923) surmised that woven textile impressions found on Neolithic (ca.
6000 to 4000 BP) pottery shards at Yangshao in Henan Province represented hemp
cloth. Li (1974b) cites many finds of Cannabis textile impressions from
China. At the early Neolithic Yang-shao site at Pan-po, near Xian in
Shaanxi Province, imprints of textiles were found on many pottery shards
dated to ca. 6225 to 5430 BP. Another Yang-shao site in Shaanxi Province
yielded pottery spinning whorls, fine bone needles and textile impressions in
the dirt of one grave and were interpreted as hemp re-mains. Bronze weapons of
the Shang Dynasty (ca. 3764 to 3120 BP) recovered from excavations at
Anyang, Henan province, have impressions left by cloth wrappers thought to be
hemp. A dagger recovered from a ca. 2900 to 2800 BP burial at Ning Xian,
Gansu Province was apparently wrapped in hemp or a similar coarse fabric as
indicated by fiber impressions (So and Bunker 1995).
By ca. 3000 BP, Cannabis
had most likely migrated west and south over the Himalayas and into India,
probably coming with nomads and traders over the trade routes that crossed the
region. In light of the accepted antiquity of Cannabis in India, it is
noteworthy that no Cannabis re-mains have been recovered from
archeological sights there.
Although archaeological and
historical data provide a foundation for our understanding of Cannabis
dispersal in Asia, there remains a severe lack of palynological and
archeological references with which to correlate these data. We were not able to
find many references dealing with analytic evidence of Cannabis pollen
for the entire Asian region and no archeological finds of Cannabis
remains at all from southern India. Certainly, archeological sites have been
investigated, but translations of foreign studies appear to be rare. This may
simply be the result of researchers focusing their investigations on other
topics besides Cannabis remains. Many early excavations overlooked
botanical evidence in their search for cultural objects. Long core samples
dating further back in time may reveal Cannabis pollen grains giving us a
much earlier time scale for the origin, evolution and migration of Cannabis.
This is an area worth pursuing and will help broaden our biological and
historical knowledge of this important crop plant.
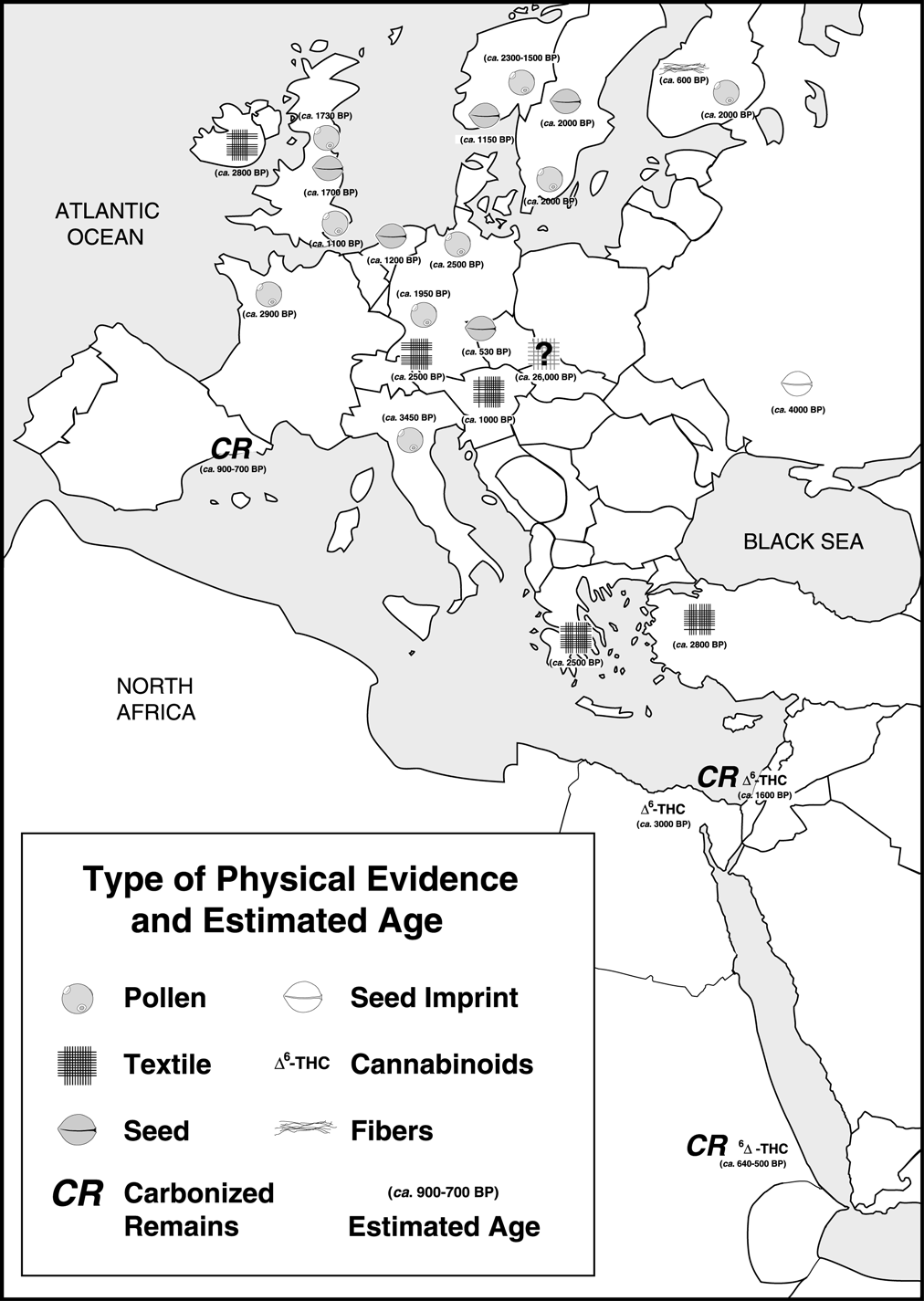
Figure 4. European Cannabis remains and approximate ages.
Evidence for the diffusion of Cannabis in Europe and
the Middle East
The most comprehensive English
language survey of the archeological evidence for Cannabis early history
in Europe is by Godwin (1967b). Dörfler (1990) provides a more recent German
language overview of both macro-fossil and pollen evidence for the early
distribution and diffusion of Cannabis in Europe.
According to the pollen record, the
spread of Cannabis throughout Europe was a two-pronged migration. After
its establishment in the Balkan states and Italy during the time of the Greek
and Roman empires, it spread both north and west through eastern and southern
Europe. Trade between the Vikings and various Mediterranean cultures resulted in
its early establishment and cultivation in Scandinavia and its subsequent spread
into the rest of northwestern Europe (Figure 4).
There is little palynological
evidence of the occurrence of cultivated Cannabis from Greek or Roman
classical sites, but its use as a foodstuff, fiber, oil, medicine, and drug is
often noted in the literature of the time. Cannabis occurrence in the
northern provinces of Italy was rare (Godwin 1967b), but Caramiello et al.
(1992) did find pollen of both Cannabis and Humulus at sites in
southern Italy dating to ca. 2500-2200 BP.
Willis (1992) studied the pollen
stratigraphy of late Quaternary deposits in northwest Greece and noted no
occurrences of Cannabis or Humulus and only minute quantities of
Cannabaceae indicator species (described below). He suggested that the region
was not suitable for agriculture and local tree clearances reflected in the
pollen record were made to support animal husbandry. This hypo-thesis seems to
agree with historical evidence for the area.
Godwin (1967b) further noted that,
"Etymological evidence seems to indicate that knowledge of the plant and its cultivation were carried to western Europe by the migrating Teutonic peoples. For the purpose of checking this assumption, we may most usefully turn to pollen analytic evidence, for whereas ropes and textiles may well have been imported, cultivation of hemp will be directly registered in the pollen rain."
Cannabis pollen values tend to be strongly associated
with other indicators of arable cultivation (Plantago, Rumex, Artemisia,
several species of the Chenopodiaceae) and often with flax (Linum), and
with high frequencies for rye (Secale) as well as certain specific tree
clearances, generally deciduous trees and sometimes Pinus. This is well
documented in pollen studies throughout continental Europe, Scandinavia and
England. Miotik-Szpiganowicz (1992) documented high Cannabaceae pollen curves in
Bory Tucholskie in northwest Poland. Taking three cores from the lake, she noted
the first Cannabis/Humulus pollen appearance at ca. 5500 BP
correlated with radio-carbon dating and varve analysis (e.g. an
examination of different layers of sediment). During this time, there was also
an increase of ash (Fraxinus) and a decrease in hazel (Corylus)
pollen deposits, certainly the results of the first Neolithic human influences
in the region. She similarly noted the elm (Ulmus) decline in the area
that she dated to ca. 5100 BP, and which has been documented throughout
all of northern Europe. A warm climate for the time period was confirmed by
significant values of mistletoe (Viscum) and ivy (Hedera) pollen
curves. (Note: These curves are not shown on her pollen diagrams, but they are
discussed in her text.) By ca. 3200 BP, millet (Setaria), barley (Hordeum),
wheat (Triticum), and rye (Secale) are all present in the diagram,
and by the end of the Roman period (ca. 1800 BP) hemp, flax, and vetch (Vicia)
are indicated. The local Cannabis maximums do not show in-disputable
evidence of cultivation until ca. 1000 BP where there is a subsequent
increase in hemp, plantain (Plantago), ragweed (Artemisia), rye,
wheat, and cereal pollen curves and a decrease in oak (Quercus), elm,
birch (Betula) and alder (Alnus) pollen. The high Cannabaceae
percentages continue into the last century, then disappear; dioecious tree
pollen is still infrequent although pine pollen increased gradually over time.
This study has been supported by
other studies in Poland and the former Czechoslovakia by Balaga (1990), Latalowa
(1992) and Ralska (1992). Balaga studied areas in southern Poland and northern
Czechoslovakia and mentioned the "invasion of the Scyths into the Lusatain"
region ca. 2000 BP. He discussed in his studies many anthropogenic
indicators of hemp cultivation, but showed no Cannabis pollen. Decreases
in oak, elm, hazel, and linden (Tilia) with corresponding increases in
plantain (Plantago), dock (Rumex), and the cereals were shown.
Latalowas study on Wolin Island in the Baltic Sea showed "much hemp
pollen occurring from 1000 BP to [the] present" with high values of winter
rye and barley, possibly indicating crop rotation, a method practiced in Asiatic
cultivation of Cannabis. Ralskas study revealed Cannabaceae pollen in
small, but constant occurrences from ca. 7000 to 3900 BP in central
Poland. This curve was interpreted to be that of Humulus, due to its
antiquity and slight occurrence.
Delusina (1991) investigated Holocene
pollen stratigraphy in Lake Ladoga near St. Petersburg, Russia in an attempt to
correlate pollen zones of the area with those of the neighboring Russian region
of northern Karelskaya and southern Finland. She found no Cannabaceae pollen,
but noted great quantities of pondweed (Potamogeton), an indicator of
many retting sites in England (Bradshaw et al. 1981).
Central, southern and western Europe
have also yielded some interesting palynological evidence for hemp and hop
occurrence. Lutgerink (1989) documented local occurrences of both Cannabis
and Humulus in east-central France. Unfortunately, the study was not
correlated with radiocarbon dating, and birch was the sole source of arboreal
pollen. This study took place in an elevation zone of 1300-1700 meters that
showed no arable cultivation. From northwestern France, Corillion and Planchas
(1963) reported high values for Cannabaceae pollen associated with rye and weeds
of arable ground from ca. 2000 BP. They furthermore note historical
accounts of widespread hemp cultivation from the 11th through the 18th
centuries. Further west in France, van Zeist (1964) showed high Cannabaceae
values also associated with rye cultivation. Radiocarbon dating showed that
continuous curve to be low from ca. 2900 BP, but with high occurrences
after ca. 2600 BP. Welten (1952) was able to date Cannabis pollen
to ca. 2200 BP in Spitzierbucht, Switzerland, but wide-spread cultivation
there occurred during the 12th through the 17th centuries. His studies showed
the Cannabis/ Humulus maximum to reach 10-13% of the total pollen,
and showed occurrences of the usual indicator species. From Sehestedt on the
northwestern German coast, Wiermann (1965) reported a substantial Humulus
curve paralleled by clearance indicators, especially those of arable ground (i.e.,
cereals, Cruciferae, and Centaurea cyanus). His studies showed
intermittent occurrences from ca. 2500 BP, continuous but low values from
ca. 1600 BP, and high values after ca. 600 BP. Hölzer and Hölzer
(1995) documented Cannabis pollen near Stuttgart, Germany from ca.
1950-250 BP.
The most recent recovery of Cannabis
pollen is from the Po Plain in northern Italy, a region famous for hemp
cultivation during the historical period. Ravazzi (pers. comm. 1998) reports Cannabis
pollen (positively identified based on pore structure and grain size) from a
Middle Bronze Age (ca. 3500-3400 BP) village site. Cannabis pollen
percentages reach 30% near the river and 8% far from the river. These layers
also contain pollen of several cultivated and weed plants.
The literature on palynological
studies from Scandinavia, Denmark, and Great Britain is overwhelming. Godwin
(1967a,b) has studied the vegetational history of the region and local
occurrences of Cannabaceae are well documented. Godwin (1967a,b) summarized the
work of several authors on the history of Cannabaceae occurring in England and
Europe, in addition to providing his own data from Old Buckenham Mere, which is
often cited. Based on his information, he concluded that hemp was not cultivated
in England before ca. 2000 BP. Indeed, his data show that hemp was not
cultivated in the area until the end of Roman times when the Anglo-Saxons
invaded the island (ca. 1500 BP), bringing with them the ox-drawn plow.
His Cannabaceae curve is mirrored by increases in Secale, Linum,
and Triticum. The increased emphasis on arable cultivation during
Anglo-Saxon and Norman times is evident, showing continuous curves for the
arable crops. Cannabis maximums appear to occur from ca. 1200-800
BP and suffer a considerable setback around the 14th century.
Bartley et al. (1976) shows Cannabis
pollen from the Durham lowlands that comprise 19% of the total pollen count in a
continuous curve from 1730 to 852 BP. Birks (1965) shows a Cannabis
occurrence from Cheshire Moss dating to ca. 1350 BP. Peglar (1993)
studied the vegetational history around Diss Mere on the eastern English coast
and revealed an almost continuous Cannabis pollen curve for the area from
ca. 1100-150 BP. Using a pollen percentage diagram, she showed the almost
mirror-like occurrence of hemp with cultivated rye, flax, barley, and mustard (Brassica)
as well as the near omnipresence of plantain and nettle (Urtica).
Historical records show the area was the center of the hemp industry in England.
Pursehouse (1961) noted that 15% of all sown areas in the Waveney valley
supported hemp, and her study correlates with this information nicely. Her
diagram showed a very rich pollen assemblage, and there are good records of the
flora and climatic conditions around the area, but her study had some potential
weaknesses. First, the sediments she studied were calcareous, which can skew
carbon-14 dating (Peglar 1993), making pollen-influx diagrams and rates of
palynological change impossible to accurately document. Secondly, she admitted
that sedimentation rates for the area may also show marked changes, and local
disturbances are likely to have re-deposited pollen grains in the study site.
A study from Kentmore in
Westmoreland, Wales by Walker (1955) sampled calcareous and detritus mud for
Cannabaceae pollen. Using Godwins (1940) British numbering system, he noted
occurrences of Cannabaceae pollen corresponding with decreases in oak, elm,
birch, alder and increases in plantain, ragweed, and Cyperaceae. He also noted a
large curve for pondweed, a retting site indicator as noted by Bradshaw et al.
(1981). Walker was unable to distinguish between hemp and hop pollen, and
attempts at verification by other scientists could not distinguish the two
genera.
A brief discussion on hops
occurrence is in order here. Wilson (1975) discussed the occurrence of Humulus
lupulus in central Europe and noted that its natural range extends
throughout most of Europe (except the islands of Crete, Iceland, and Spitzbergen)
over the Caucasus and Altai Mountains into Siberia, north up to the Arctic
Circle and south into Morocco and Palestine. Based on this early evidence,
Wilson concluded that "Cannabaceae pollen curves from Zone VIIb and earlier
in northern Europe are assumed to be that of hop only," using the
occurrence of Cannabis grains as post-Roman indicators. The first written
evidence of hop cultivation in Europe was ca. 1140 BP (DeLyser and Kasper
1994). Increases of Cannabaceaen pollen curves could not be attributed to
increases in hop cultivation until after 1100 BP.
Cannabaceae pollen grains and Cannabis
seeds were observed by Bradshaw et al. (1981) in Flandrian deposits in
eastern England. His studies showed the pollen curve to reach up to 50% of the
total land pollen in places and similarly noticed much pondweed pollen and many
molluscan shell fragments, indicating that his samples came from sediments that
once stood underwater. His pollen diagram also showed a drying-out period
evidenced by a decrease in Cannabaceae pollen and local maxima of Equisetum,
Sphagnum, and Cyperaceae pollen. Evidence from his earlier studies
in other areas revealed that Cannabaceae maximums in those samples never
exceeded 25% and lacked shell fragments and pondweed grains. Bradshaw concluded
that this site was a retting site, but could not decide if local hemp
cultivation had occurred or not.
French and Moore (1986) refuted
Bradshaws hypo-thesis with an independent study from Cors Llyn in central
Wales. Historical evidence documents the cultivation of Cannabis around
the lake, first on the eastern side, later on the western side. Their pollen
data showed extensive Cannabis cultivation around the area and indicate
that retting did not occur at this site based on the following facts: (1)
retting involves considerable disturbance of a site, both of sediments and
pollen profile and neither disturbance was observed at the study area; (2) Cannabis
pollen was still observed after the beginning of a schwingmoor (boggy peat land)
formation and the lake would be unsuitable for retting at this stage; (3) if
retting had occurred, one would expect a substantial Cannabis curve like
that seen in Bradshaws study, but the pollen percentages reported by French
and Moore (1986) never exceeded 12% of the total land pollen; (4) an
agricultural survey of southern Wales in 1815 indicated that the region was damp
enough to allow for retting by simply laying the hemp plants outside over the
winter; (5) a calculation of diversity of pollen types from the pollen diagram
indicated an increase of pollen-type diversity during the Cannabis
stages, not a decrease, as retting would reveal. Further studies, and a
re-examination of previous palynological studies using the criteria mentioned by
French and Moore (1986), will help to give more detail to the emerging picture
of hemp cultivation and processing.
Other evidence for Cannabis/Humulus
comes from Scandinavia and Denmark. These data help make clear that the Vikings
were responsible for the introduction of hemp into northwestern Europe. Hafsten
(1956) showed data from the inner Oslo Fjord area of Norway that indicate the
probable occurrence of cultivated hemp from ca. 2300-1500 BP. His pollen
curves showed maxima for Cannabaceae to be 45% in some spots, but later evidence
from Bradshaw et al. (1981) indicate that this high percentage may be
due to retting (as previously discussed) and we therefore must also note that
possibly, the Vikings could have been introducing foreign-grown hemp for
retting, although, hemp is much easier to transport after it is retted and
peeled from the stalks.
In Sweden, Fries (1962) has done
pollen studies near Osbysjo and shown a Cannabis/Humulus type
pollen curve that begins with low occurrences ca. 1850 BP and reaches 13%
during the Viking period of ca. 1150-800 BP. It should be noted that his
curve was not correlated with other arable cultivars, but a curve for rye does
appear from ca. 2000 BP with high values after ca. 900 BP. Lageras
(1995) showed Cannabis pollen from multiple sites in southern Sweden from
ca. 600-200 BP. Godwin (1967a) found Cannabis pollen in
southeastern Sweden, associated with rye pollen, dated from ca. 1600-800
BP. Tolonen (1978) showed Cannabaceae pollen curves from southern Finland. He
interpreted Humulus pollen from ca. 3530-2639 BP and Cannabis
pollen from ca. 2000-500 BP. Studies done by Anderson (1954) in southern
Jutland show a significant Cannabaceae maximum dated to ca. 400 BP.
However, as Cannabis is easily confused with Humulus pollen, Cannabis
macrofossils (e.g., fibers, seeds, trichomes) may prove to be more
definitive than, or at least a valuable supplement to, pollen evidence.
Early European and Middle Eastern
finds of Cannabis fibers are much less common than in China. Barber
(1991) reports that hemp and linen fibers were found snagged in a bone tool
recovered from a late Neolithic (ca. 4000 BP) site at Adaouste in
southern France and also mentions the recovery of hemp fabric scraps from the
ruins of Gordium, Turkey dated at ca. 2800 BP and Trakhones, in the
Attiki Province of Greece dated at ca. 2500 BP. None of these
fiber analyses have been substantiated. Brown layers containing Cannabis
fibers found on lake bottoms in Finland have been interpreted as indicating the
soaking of hemp after ca. 600 BP (Saarnisto et al. 1977). Cannabis
fibers recovered from lake sediments provide more certain evidence of local hemp
retting than that provided by pollen grains.
Hemp string and fabric specimens were
recovered from a Bronze Age (ca. 2800 BP) site at St. Andrews in Scotland
(Ryder 193). Identification was based on measurements of fiber diameter and
comparisons were made with flax samples. Delaney (1986) refers to a Celtic
"princely tomb" dated from the late Hallstatt period (ca. 2550
to 2500 BP) in Hochdorf, Germany excavated in 1978-1979 by Jorg Biel,
"Here the people of prehistoric Hochdorf buried a Celtic chieftain who merited a great mausoleum. He lay on a bronze, high-backed couch embossed with ritual dancing figures and horses pulling a cart. . . . Dr. Biels fiber analysis revealed, imbedded in the bronze, horsehair, hemp, wool, and the fur of badgers, on which the dead prince had reposed."
Textile fragments
from an early Magyar graveyard (ca. 950 BP) at Halimba-Cseres in Hungary
were made of either hemp or flax (Török 1954). The hemp fibers were
differentiated by microscopical observation.
In continental Europe, seed remains
have been report-ed from two pre-Roman Iron Age (ca. 3000 to 2500 BP)
sites (Willerding 1979), a water well dated at ca. 530 BP in the Bohemian
region of Czechoslovakia (Opravil 1979), and many sites dating from ca.
1200 - 350 BP in the Netherlands (Pals pers. comm. 1997). In Britain, Cannabis
seeds have been recovered from a Roman era (ca. 1800 to 1600 BP) well
(Figure 2) at Skeldergate (Hall et al. 1980) and Medieval pits and
watercourses at Sewer Lane, both in York (Williams 1977). Scandinavian seed
remains have been recovered from a few sites in Sweden dating from as early as ca.
2000 BP (Påhlsson 1982 and Gaillard pers. comm. 1997). Although no hemp ropes
or textiles were reported from the famous Oseberg ship burial of a woman (ca.
1150 BP), four Cannabis seeds were recovered (Holmboe 1927) and are now
believed to be connected with the womans priestly functions (Christensen
1992).
Fruits of Cannabis have also
been found in Viking settlements in Denmark (Godwin 1967a,b). Rather
surprisingly, there is no solid historical or etymological evidence that hemp
was known to Iceland until quite late in medieval times, first noted in the Kornungs
Skuggsja written in ca. 1240 AD (Godwin 1967b). As Godwin (1967b)
points out,
"The historical evidence as a whole clearly points to cultivation of hemp in the Middle East...It spread rapidly in the Mediterranean area in classical times but there is very little evidence that its cultivation was extended northwards within the Roman empire".
Seed impressions
attributed to Cannabis have been found in several clay shards recovered
from one middle Neolithic (ca. 5000 BP) site of the Linearbandkeramik
Culture north of the Black Sea. Imprints of peas (Pisum sativum) and a
vetch (Lathyrus sp.) were also found. Some charred remains or imprints of
Cannabis seeds are also associated with Iron Age (ca. 2800 to 2400
BP) Scythian remains from the Ukraine (Yanushevich 1989).
Israeli researchers discovered
carbonized material in the tomb of a 14 year old girl excavated near Jerusalem
and dated at 1600 BP. Initial microscopic investigation indicated that the
material possibly resulted from burning a mixture of Cannabis and other
plants. Subsequent chemical analysis revealed the presence of Δ6-THC.
Researchers concluded that Cannabis had been burned to facilitate the
birth process (Zias et al. 1993).
Evidence for the diffusion of Cannabis in Africa
Cannabis was introduced
into Africa at an early date, although accounts of this differ greatly and no
certain date has been agreed upon. Schultes (1970) estimates that the period of ca.
4000-3000 BP saw the first introduction of Cannabis into Africa, but the
first physical evidence that the plant reached the African continent is not
given until the ca. 640-500 BP (van der Merwe 1975). Whichever the case,
its use in Morocco for drugs was firmly established by the early 1800s
(Clarke 1998). The only pollen record we were able to locate from Africa was
modern pollen trapping in Durban, South Africa (Cadman and Dames 1993). The
authors tested aerial pollen for three years in the late 1980s and Cannabis
pollen accounted for an average of 20% of the total pollen count over the test
period. The authors concluded that this count indicated local cultivation.
Pipes thought to have been used for
smoking Cannabis have been recovered from both Ethiopia and southern
Spain. Remains of two ceramic water pipe bowls excavated from Lalibela cave in
Ethiopia were dated at ca. 640 - 500 BP. Both contained trace amounts of
³6-THC. This evidence was interpreted to indicate that psychoactive Cannabis
containing THC was smoked in these pipes (van der Merwe 1975). Juan i Tresserras
(pers. comm. 1998) claims to have identified charred Cannabis remains (i.e.,
parenchyma tissues, siliceous hairs and calcium oxalate crystals) from an
Arabian pipe recovered from the medieval Christian castle site of Cornellà de
Llobregat, Barcelona and dated at ca. 900-700 BP. The methods used to
identify the charred remains as Cannabis were not disclosed. These
reports are controversial, as both these dates precede the exploration of the
New World by Spain and the supposed first date of introduction (ca. 400
BP) of tobacco, pipes and smoking from the New World into Europe. We hope for
more substantive data in the near future.
Additional archeological evidence
attributed to Cannabis use were found in the body tissues of several
Egyptian mummies chemically determined to contain cannabinoids (Balabanova et
al. 1992). According to their published reports,
"The immunological determination of cocaine and hashish (THC) showed the presence of these drugs in all nine samples."
"This is the first study which shows the presence of cocaine, hashish and nicotine in Egyptian mummies, dating back to about 1000 BC. This means that these three organic substances are capable of surviving in hair, soft tissue and bones for ca. 3000 years under favorable conditions. However, it cannot be deter-mined at present whether the concentrations measured represent the original amount of these drugs during life or immediately after death, or what kind of decomposition might have taken place in the past 3000 years."
This is the only
archeological study linking Cannabis with ancient Egypt. Historically,
the ancient Egyptians used flax (Linum) and cotton (Gossypium) for
spinning and weaving and there is no mention of Cannabis use for any
purpose. The presence of cannabinoids in the tissues of Egyptian mummies brings
up the possibility that Cannabis was used recreationally/religiously or
medicinally by the early Egyptians. However, most of the controversy centers
around the reports of cocaine and nicotine contents in these Egyptian mummies
predating Columbus "discovery" of the New World. The plant genera Erythroxylum
(the sole source of cocaine) and Nicotiana (the sole source of nicotine)
are both considered to have only a New World distribution prior to European
contact during the 15th century, much later than the dates (ca. 3000 BP)
of the mummies analyzed by Balabanova et al. (1992). These results are so
unusual that they cast some doubt over the cannabinoid findings as well.
Claims from the same research team
add even more mystery. According to Wills (1998),
"In 1993, a team of German anthropologists published the results of an analysis of various tissues from 72 Peruvian mummies dated ca. 1800-500 BC (Parsche et al. 1993). Bones from 20 of them were shown to contain cannabinoids. In the same study, ten bodies from the German Bell Culture (ca. 4500 BP) did not contain cannabinoids. In addition, two African mummies from the Sudan (dated at ca. 7000 to 6000 BP) and 1600 to 600 BP) also did not contain Cannabis."
Diffusion of hemp in the New World
Haney and Kutscheid (1973) documented
the spread of Cannabis from the east coast westward into Missouri (1835),
Illinois (1875), Nebraska (1887), California (1912) and Michigan (1922), and van
Zant et al., (1979) studied Cannabis-bearing deposits at Lake West
Okoboji, Iowa. The rise of Cannabaceae pollen in this study is presumed to
reflect the planting of hemp crops there in 1910. MacQuiddy (1995) showed
significant pollen counts near Omaha, Nebraska in air trapping studies of pollen
there, and Basset et al. (1978) caught large amounts of Cannabis
pollen on exposed slides in Ottawa, Canada. Although these last two studies were
not of pollen cores taken from the soil, they may be useful in that they provide
a comparative baseline of data to use in studies of past Cannabis
diffusion.
Conclusion
The positive identification of the
physical remains of hemp textiles, cordage, pollen grains, fruits, stalks and
trichomes, combined with chemical analysis for trace remains of cannabinoids are
techniques used to establish a prehistoric distribution for Cannabis.
Palynological and other archaeological evidence substantiate much of the
historical evidence for the origin of Cannabis in Asia and its migration
around the globe. Further study is needed to clear up the lack of corroborating
physical data for many regions.
References
Anderson, A. 1954. Two standard diagrams from south Jutland. Danmarks Geol. Untersolgelse II 80: 188-209.
Andersson, J. G. 1923. An early Chinese culture. Bull. Geog. Survey China 5(1): 26.
Anonymous 1994. Scientists bid to save ancient princess. China Daily 14(4126): 1.
Bahn, P. G. 1996. Tombs, Graves and Mummies. Weidenfeld & Nicholson, London: 96, 106, 146, 164.
Balabanova, S., F. Parsche and W. Pirsig 1992. First identification of drugs in Egyptian mummies. Naturwissenschaften 79: 358.
Balaga, K. 1990. The development of Lake Lukcze and changes in the plant cover of the south-western part of the Lenczna-Wlodawa lake district in the last 13000 years. Acta Palaeobot. 30(1,2): 77-146.
Barber, E. J. W. 1991. Prehistoric Textiles - The development of cloth in the Neolithic and Bronze Ages with special reference to the Aegean. Princeton University Press, Princeton: 16-18.
Basset, I. J., C. W. Crompton and J. A. Parmalee 1978. An Atlas of Airborne pollen grains and common fungus spores in Canada. Monograph 18. Canada Dept. of Agriculture, Ottawa: 321 pgs.
Birks, H. J. B. 1965. Pollen analytical investigations at Holcroft Moss, Lancashire and Lindlow Moss, Cheshire. Journal of Ecology 53(2): 299-314.
Bradshaw, R. H. W. et al. 1981. New fossil evidence for the past cultivation and processing of hemp (Cannabis sativa L.) in eastern England. New Phytol. 89: 503-510.
Cadman, A. and J. F. Dames 1993. Airspora of Durban: a sub-tropical, coastal South African city. I. Pollen component. Grana 32(6): 372-375.
Caramiello, R. et al. 1993. Palynological findings in the Centocamare and Marasà Sud sites (Locri Epizephyrii) and their relation to the historical and archeological hypotheses. Alloionia 31: 7-19. [in Italian]
Chang, K. 1963. The Archeology of Ancient China. Yale Univ. Press, New Haven and London: 120, 173.
Cheng, T.-K. 1959 Archeology in China, Vol. 1 - prehistoric China. W. Heffer and Sons Ltd., Cambridge: 11-168.
Cheng, T.-K. 1963 Archeology in China, Vol. 3 - Chou China. W. Heffer and Sons Ltd., Cambridge: 36-37.
Chou, K. 1963. Analysis of pollen from the Neolithic site at Pan-po. App. 3, Si-an Pan-po. Peking.
Christensen, I. M. 1992. Osebergdronningens gravvaar arkeologiske nasjonalsskatt i nytt lys. Oslo, Schibsted. [in Norwegian]
Chrtek, J. 1981. Bemerkungen zur verbreitung der art Cannabis ruderalis in Böhmen und Mähren [Remarks on the distribution of the species Cannabis ruderalis in Bohemia Czechoslovakia.] Casopsis Narodniho Muzea 150(1,2): 21-24. [in German]
Clarke, R. C. 1995. Scythian Cannabis verification project. Journal of the International Hemp Association 2(2): 104.
Clarke, R. C. 1998. Hashish!. Red Eye Press, Los Angeles: 387.
Corillion, R. and N. Planchais 1963. Recherches sur la vegetation actuelle et passee dune lande torbeuse Armoricaine: Malingue (Meyenne). Pollen Spores 5: 273-286.
Delaney, F. 1986. The Celts BBC Publications, London: 19, 22.
Delusina, I. V. 1991. The Holocene pollen stratigraphy of Lake Ladoga and the vegetational history of the surroundings. Ann. Acad. Sci. Fennicae 153(3): 1-66.
DeLyser, D. Y. and W. J. Kasper 1994. Hopped beer: The case for cultivation. Economic Botany 48(2):166-170.
Dörfler, Walter 1990 Die geschichte des hanfanbaus in Mitteleuropa aufgrund palynologischer untersuchungen und von Großrestnach-weisen Praehistorische Zeitschrift 65:218-244. [in German]
French, C. N. and Moore, P. D. 1986. Deforestation, Cannabis cultivation, and schwingmoor formation at Cors Llyn (Llyn Mire), central Wales. New Phytol. 102: 469-482.
Fries, M. 1962. Studies of the sediments and the vegetational history in the Osbysjo Basin, north of Stockholm. Oikos 13: 76-96.
Gaillard, M.-J. 1997. personal communication.
Garg, A. 1996. Palynocontents of bee-collected pollen loads of autumn season in Bhimal, India. Taiwania 41(3): 197-207.
Godwin, H. 1940. Pollen analysis and forest history of England and Wales. New Phytol. 39: 370.
Godwin, H. 1967a. Pollen-analytic evidence for the cultivation of Cannabis in England. Rev. Palaeobot. Palynol. 4: 71-80.
Godwin, H. 1967b. The ancient cultivation of hemp. Antiquity 42: 42-49.
Grudzinskaya, I. A. 1988. On the taxonomy of Cannabaceae. Bot. Zhurn. 73(4): 589-593.
Hafsten, U. 1956. Pollen analysis investigations on the Late-Quaternary development in the inner Oslo Fjord area. Arbok. Univ. Bergen, Mat. Naturv. Ser. 8: 1-162.
Hall, A. R., H. K. Kenward and D. Williams 1980. Environmental evidence from Roman deposits at Skeldergate. in Addyman, P. V. (Ed.) The Archeology of York 14(3):101-156.
Haney, A. and B. B. Kutscheid 1973. Quantitative variation in the chemical constituents of marihuana from stands of naturalized Cannabis sativa L. in east central Illinois. Econ. Bot. 27: 193-203.
Harlan, J. R. 1986. Plant domestication: diffuse origins and diffusions. in Barigozzi, C. (Ed.) The Origin and Early Spread of Agriculture in the Old World. New York: Elsevier.
Holmboe, J. 1927. Osebergfundet. in Den norske. Stat. Oslo. [in Norwegian]
Hölzer, A. and A. Hölzer 1995. Studies on the vegetation history of the Hornisgrinde area in the northern Black Forest (SW Germany) by means of pollen, macrofossils and geochemistry. Carolinea 53: 199-238. [in German]
Juan i Tresserras, J. 1998. Personal communication.
Kansu Museum 1972. Preliminary report on the excavation of 3 Han tombs at Ma-tsu-tze, Wu-wei. Wen-wu 12: 9-21.
Kuo, Y. 1978. The Western Han wooden-chambered tomb of Lao-fu-shan, Nan-chang, Kiangsi province KK 1965/66: 268-272, 300, pl. II, III in Rudolph, R. C. (Ed.) Chinese Archeological Abstracts. UCLA Inst. of Archeology, Los Angeles: 252.
Joseph, R. 1973. Economic significance of Cannabis sativa in the Moroccan Rif. Econ. Bot. 27: 235-240.
Lagerås, P. 1996. Long-term history of land-use and vegetation at Femtingagölen - a small lake in the Småland Uplands, southern Sweden. Veget. Hist. Archeobot. 5: 215-228.
Latalowa, M. 1992. Man and vegetation in the pollen diagrams from Wolin Island (NW Poland). Acta Palaeobot. 32(1): 123-249.
Lewis, W. H., V. Prathiba and V. E. Zenger 1983. Airborne and allergenic pollen of North America. Johns Hopkins Univ. Press, Baltimore: 150-153.
Li, H. L. 1973. The origin and use of Cannabis in East Asia: linguistic and cultural implications. Econ. Bot. 28: 293-301.
Liou, S. Z. 1988. Moraceae. in Li, S. X. (Ed.) Flora Liaoningica 1: 281-292.
Li, H. L. 1974. An archaeological and historical account of Cannabis in China. Econ. Bot. 28(4): 437-448.
Lutgerink, C. A. et al. 1989. Regional pollen assemblages versus landscape regions in the Monts du Forez, Massif Central, France. Pollen et Spores 31(1,2): 45-60.
MacQuiddy, E. L. 1955. Northern prairies and plains (Iowa, Nebraska and the Dakotas) in Samter, M. and O. C. Durham (Eds.) Regional Allergy of the United States, Canada, Mexico and Cuba. C. C. Thomas, Springfield, Illinois: 183-195
Miotik-Szpiganowicz, G. 1992. The history of the vegetation of Bory Tucholskie and the role of man in the light of palynological investigations. Acta Palaeobot. 32(1): 39-122.
Nordal, A. 1970. Microscopic detection of Cannabis in the pure state and in semi-combusted residues. in Joyce, C. R. B. and S. H. Curry (Eds.) The Botany and Chemistry of Cannabis. J. & A. Churchill, London: 61-68.
Opravil, E. 1979. Hedera helix L. aus der mittelalterlichen Stadt Most (Tschechoslowakei). in Ludwig, M. (Ed.) Veröffentlich mit Mittelen des Land-schaftsverbandes Rheinland. Rheinisches Landesmuseum, Bonn: 209-215.
Påhlsson, I. 1982. Cannabis sativa in Dalarna. Striae 14: 79-82.
Parsche, F., S. Balabanova and W. Pirsig 1993 Drugs in ancient populations (letter). Lancet 341: 503.
Peglar, S. M. 1993. The development of the cultural landscape around Diss Mere, Norfolk, UK during the past 7000 years. Rev. Paleobot. and Palynol. 76: 1-43.
Polosmak, N. 1996. A mummy unearthed from the pastures of heaven. National Geographic Sept.: 82.
Pringle, H. 1997. Ice age communities may be earliest known net hunters. Science 277: 1203-1204.
Pursehouse, E. Hemp: A forgotten Norfolk crop. Eastern Daily Press. 25 Feb. 1961.
Ralska-Jasiewiczowa, M. and B. van Geel 1992. Early human disturbance of the natural environment recorded in annually laminated sediments of Lake Gosciaz, central Poland. Veget. Hist. Archeobot. 1: 33-42.
Ravazzi, C. 1998. (personal communication)
Rudenko, Sergei I. 1970. Frozen Tombs of Siberia - the Pazyryk burials of Iron Age horsemen University of California Press, Berkeley and Los Angeles: 34-36, 62, 197, 199-200, 284-285, Plates 35, 61c, 62, 63.
Ryder, M. L. 1993. Probable hemp fibre in Bronze Age Scotland. Archeological Textiles Newsletter 17: 10-13.
Saarnisto, M., P. Huttenen and K. Tolonen 1977. Annual lamination of sediments in Lake Lovojärvi, southern Finland, during the past 600 years. Ann. Bot. Fennici 14(1): 35-45.
Schultes, R. E. 1970. Random thoughts and queries on the botany of Cannabis. In: Joyce, C. R. B. and S. H. Curry (eds.) The Botany and Chemistry of Cannabis. London: 11-38.
Schultes, R. E. and A. Hofmann 1992. Plants of the Gods: sacred, healing and hallucinogenic properties. Rochester, VT: Healing Arts Press: 92-102.
Simpson, B. B. and M. Conner-Ogorzaly 1986. Economic Botany: plants in our world. San Francisco: McGraw-Hill: 385-391.
Small, E. 1978. A numerical and nomenclatural analysis of morpho-geographic taxa of Humulus. Systematic Botany 3(1): 37-76.
Small, E. and A. Cronquist 1976. A practical and natural taxonomy for Cannabis. Taxon 25(4): 405-435.
So, J. F. and E. C. Bunker 1995 Traders and Raiders on Chinas Northern Frontier Arthur M. Sackler Gallery, Smithsonian Institution, University of Washington Press, Seattle and London: 126.
Soják, J. 1980. Fragmenta phytotaxonomica et nomenclatorica (I.) Casopis Narodniho Muzea 148(2): 77-80.
Temple, Robert K. 1986. China- Land of discovery and invention. Patrick Stevens, Wellingborough, UK: 81-82.
Tolonen, M. 1978. Paleoecology of annually laminated sediments in Lake Ahvenainen, S. Finland. I. Pollen and charcoal analyses and their relation to human impact. Annales Botanici Fennici 15: 177-208.
Török, G. 1954 Graveyard of Halimba-Cseres dating from the X-XIIth centuries. Folia Archaelogica 4: 95-105, 207-208. [in Hungarian]
van der Merwe, N. J. 1975. Cannabis smoking in 13th-14th century Ethiopia: Chemical evidence in Rubin, V. (Ed.) Cannabis and Culture. Mouton, The Hague: 77-80.
van Zant, K. L., et al. 1979. Increased Cannabis and Humulus pollen, an indicator of European agriculture in Iowa. Palynology 3: 227-233.
van Zeist, W. 1964. A paleobotanical study of some bogs in western Brittany (Finistere), France. Palaeohist. 10: 157-180.
Vavilov, N. I. 1987. Origin and Geography of Cultivated Plants. Cambridge University Press, Cambridge, 1992. [English translation].
Walker, D. 1955. Studies in the post-glacial history of British vegetation. XIV: Skelsmergh Tarn and Kentmore, Westmoreland. New Phytol. 54(2): 222-254.
Welten, M. 1952. Uber die spat- und postglaciale Vegetationsgeschichte des Simmentals. Veroffentl. Geobotan. Inst. Rubel, Zurich 26: 1-135. [in German]
Whittington, G. and K. J. Edwards 1989. Problems in the interpretation of Cannabaceae pollen in the stratigraphic record. Pollen et Spores 31(1,2): 79-96.
Whittington, G. and A. D. Gordon 1987. The differentiation of the pollen of Cannabis sativa L. from that of Humulus lupulus L. Pollen et Spores 29(1): 111-120.
Wills, S. 1998 Cannabis use and abuse by man: An historical perspective. In: Brown, P. T. (Ed.) Cannabis - The genus Cannabis. Harwood Academic Pubs., The Netherlands: 1-27.
Wiermann, R. 1965. Moorkundliche and vegetations geschichtliche Betrach-tungen zum Aussendeichsmoor bei Sehestedt (Jadebusen). Ber. Deut. Botan. Ges. 78: 269-278.
Willerding, U. 1979. Zum Ackerbau in der jüngeren vorrömischen Eisenzeit. in Ludwig, M. Veröffentlicht mit Mittelen des Landschafts-verbandes Rheinland. Rheinisches Landesmuseum, Bonn: 309-330. [in German]
Williams, D. 1977. The plant macrofossil contents of medieval pits at Sewer Lane, Hull. in Armstrong, P. Excavations in Sewer Lane, Hull, 1974. East Riding Archeologist 3, Old Town Report Series No. 1: 18-32.
Willis, K. S. 1992. The late Quaternary vegetational history of northwest Greece. I. Lake Gramousti. New Phytol. 121: 101-117.
Wilson, D. G. 1975. Plant remains from the Graveney boat and the early history of Humulus lupulus L. in western Europe. New Phytol. 75: 627-648.
Yanushevich, Z. V. 1989. Agricultural evolution north of the Black Sea from the Neolithic to the Iron Age. in Harris, D. R. and G. C. Hillman (Eds.) Foraging and Farming - The evolution of plant exploitation. Unwin Hyman, London: 611, 614, 646.
Yin, T. 1978. The bronzes unearthed at Shu-cheng, Anhwei. KK 1964/10: 490-503, pl. I, VI in Rudolph, R. C. (Ed.) Chinese Archeological Abstracts UCLA Institute of Archeology, Los Angeles: 116.
|
|
|
|||||||||||||||||||||